

-
多囊卵巢综合征(polycystic ovary syndrome,PCOS)是育龄期女性最常见的一种生殖内分泌疾病,其特征以生化或临床高雄激素血症、无排卵和卵巢多囊样改变等为主要表现[1-2]。流行病学显示PCOS在全世界范围内的总体发病率约为6%~20%[3],而由PCOS引发的排卵障碍所致的不孕症占40%[4]。PCOS不仅影响女性的正常生殖功能,还会导致女性发生代谢系统方面的障碍,如高胰岛素血症、胰岛素抵抗,增加女性继发糖尿病、心血管疾病及子宫内膜癌的风险,对女性健康造成严重不良影响[2]。肥胖是PCOS发生的重要危险因素之一[5]。研究数据表明,28.3%的超重或肥胖的女性患有PCOS[6],多达42%的PCOS患者超重或肥胖[7]。肥胖对PCOS的发展和进展产生显著影响,研究发现,脂肪细胞主要通过分泌脂肪因子,如IL-1、IL-6、瘦素、脂联素等,作用于相应的靶器官、靶组织、靶细胞,如卵巢、肾上腺等,刺激机体产生较多的雄激素,而雄激素又可通过抑制肾上腺素受体等导致体内脂肪分解减少,脂肪大量堆积在体内,导致体内高雄激素水平与肥胖之间形成恶性循环,严重影响PCOS患者的健康状况[3]。与普通女性相比,PCOS患者具有更高的肥胖倾向,且更容易出现腹部脂肪堆积[8];而这种由于腹部脂肪堆积造成的中心性肥胖反过来又可加重PCOS患者的临床或生化表现,导致胰岛素抵抗、高雄激素血症、生殖功能异常等[9]。临床试验结果显示,若患者体质量减轻初始体质量的5%,其体内激素水平、血糖水平得以改善,同时,月经周期和排卵情况趋于正常化,这表明体质量减轻可增加患者排卵和妊娠的可能性[10]。
青蒿素(artemisinin,ART)是一种天然倍半萜内酯化合物,最初由2015年诺贝尔生理学或医学奖获得者屠呦呦从青蒿植物中提取出来并广泛用于抗疟疾治疗[11];青蒿素还用于抗癌、抗炎药物等[12]。近年研究发现,青蒿素及其衍生物还具有预防肥胖的功效:在啮齿动物模型中,青蒿素及其衍生物通过调节p38MAPK/ATF2轴和Akt/mTOR途径等在脂肪生成过程中诱导脂肪细胞褐变,从而预防肥胖并改善肥胖相关的代谢紊乱[13]。Lee等[14]和Jang[15]体外实验数据表明,青蒿素及其衍生物可通过PPARγ途径抑制脂肪生成和脂肪因子的表达。本研究通过网络药理学方法和分子对接方法分析预测青蒿素可用于治疗PCOS的潜在靶点,旨在为深入研究其治疗的作用机制提供参考。
-
通过Pubchem数据库[16](
https://pubchem.ncbi.nlm.nih.gov/ )获得天然产物青蒿素的SMILES号,并将其输入Swiss TargetPrediction数据库[17](http://www.swisstargetprediction.ch/ )进行靶点预测,导出分析结果并保存;结合PharmMapper数据库[18](http://www.lilab-ecust.cn/pharmmapper/ )预测的靶点,二者共同作为青蒿素的药物靶点,并将靶点导入Uniprot数据库[19](https://www.uniprot.org/)进行靶点蛋白与基因名称转换。以“polycystic ovary syndrome”作为关键词检索,通过DisGeNET数据库[20](https://www.disgenet.org/ )、GeneCard数据库[21](https://www.genecards.org/ )进行疾病靶点预测。将搜集的青蒿素靶点和PCOS靶点分别导入Venny在线作图软件(https://bioinfogp.cnb.csic.es/tools/venny/ )绘制韦恩图,从而得到二者的共同靶点。 -
将共同靶点导入STRING数据库[22](
https://cn.string-db.org/ ),物种选择“homo sapiens”,最低相互作用分数设置为“0.9”,隐藏游离点,其他保持默认设置,得到蛋白相互作用网络图(protein-protein interaction,PPI),将PPI网络图导入Cytoscape 3.9.1软件[23],进行核心靶点筛选。 -
将共同靶点导入DAVID数据库[24](
https://david.ncifcrf.gov/ ),分别进行基因本体(gene ontology,GO)功能、京都基因与基因组百科全书(kyotoencyclopodia of genes and genomes,KEGG)通路分析,其中GO功能富集内容从分子功能(molecular function,MF)、生物学过程(biological process,BP)、细胞组分(cellular component,CC)三部分进行逐一分析,并利用微生信在线作图软件(http://www.bioinformatics.com.cn/ )将分析结果进行可视化。 -
将所获得的青蒿素、PCOS作用靶点及信号通路分别导入Cytoscape软件构建药物-疾病-靶点-通路网络图。
-
从Pubchem数据库中下载青蒿素的2D结构,在RCSB PDB数据库[25](
https://www.rcsb.org/ )中下载核心靶蛋白结构。利用Chem3D软件对青蒿素的2D结构进行转化,用Pymol软件对核心靶蛋白结构进行初步处理,再用Auto Dock Tools软件做进一步加氢等处理,并将处理的核心靶蛋白保存为“pdbqt”格式进行分子对接,最后利用Pymol软件对分子对接结果进行可视化处理[26-27]。 -
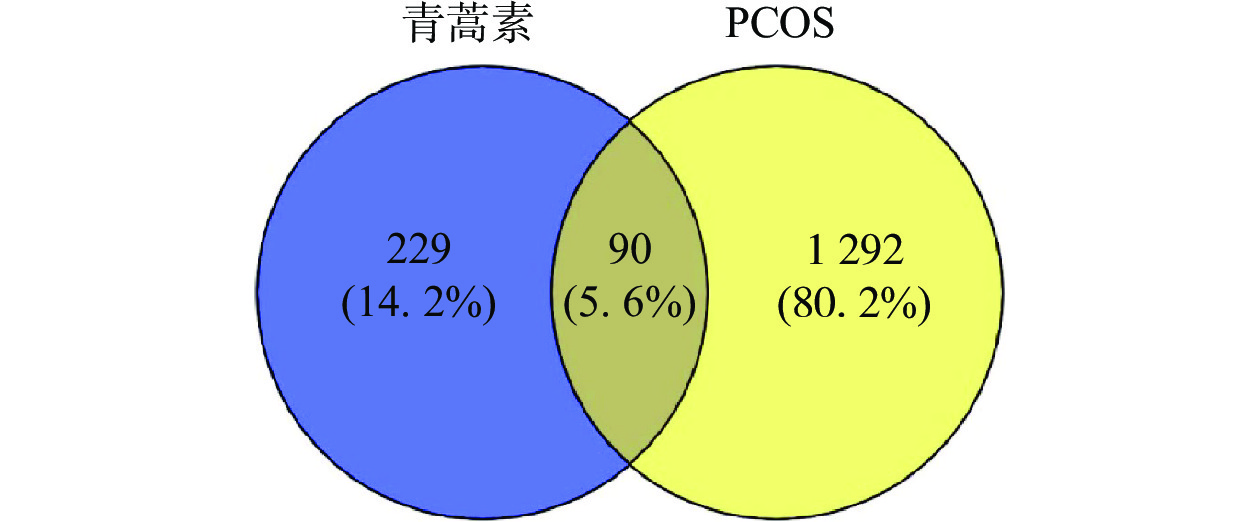
通过数据库检索共得到青蒿素潜在作用靶点229个,PCOS疾病靶点1292个。利用在线作图软件将青蒿素作用靶点与PCOS疾病靶点进行韦恩图分析,得到二者的交集靶点90个,如图1所示。
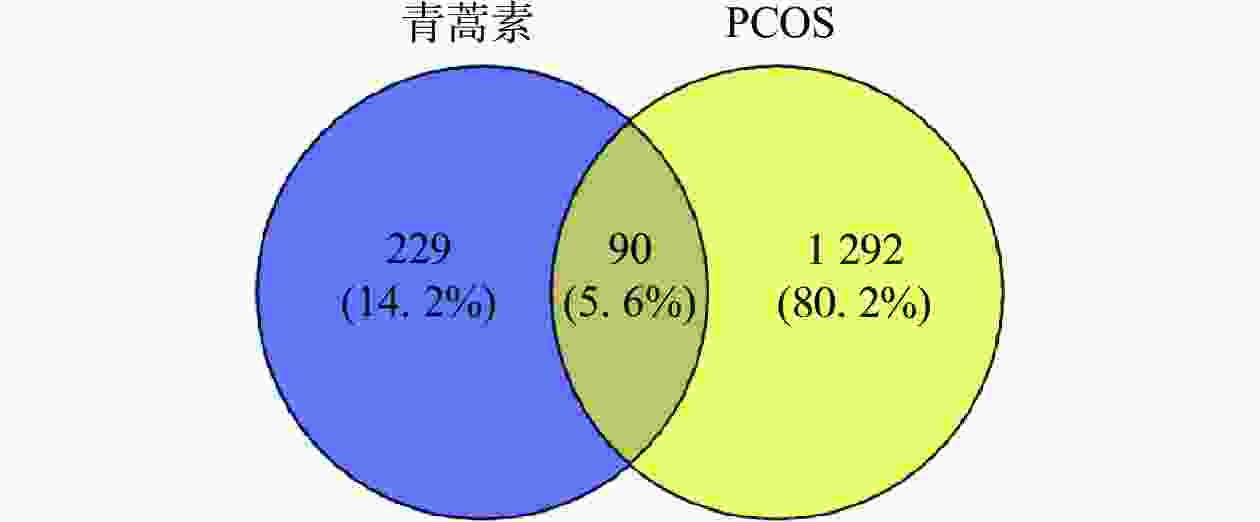
-
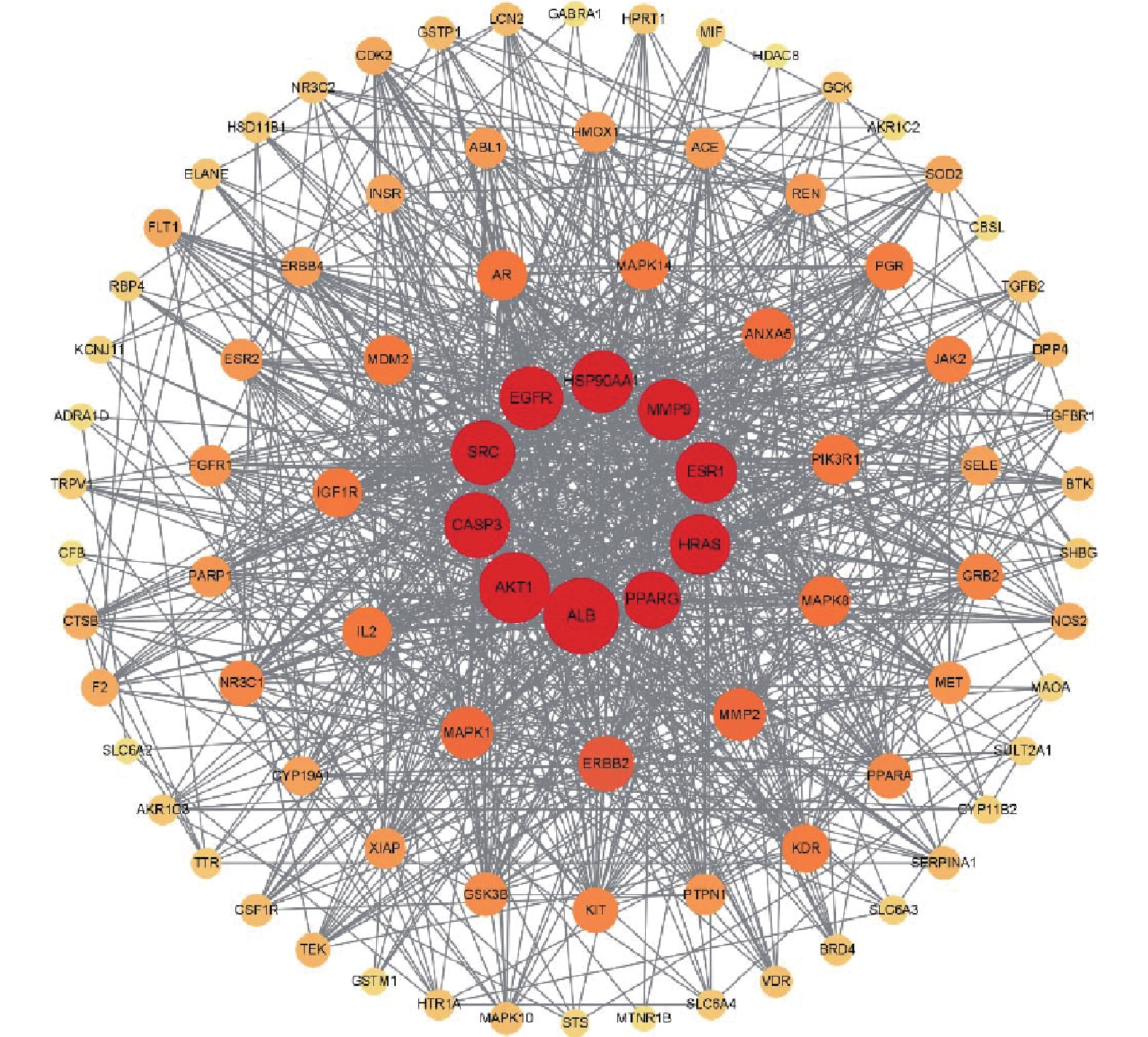
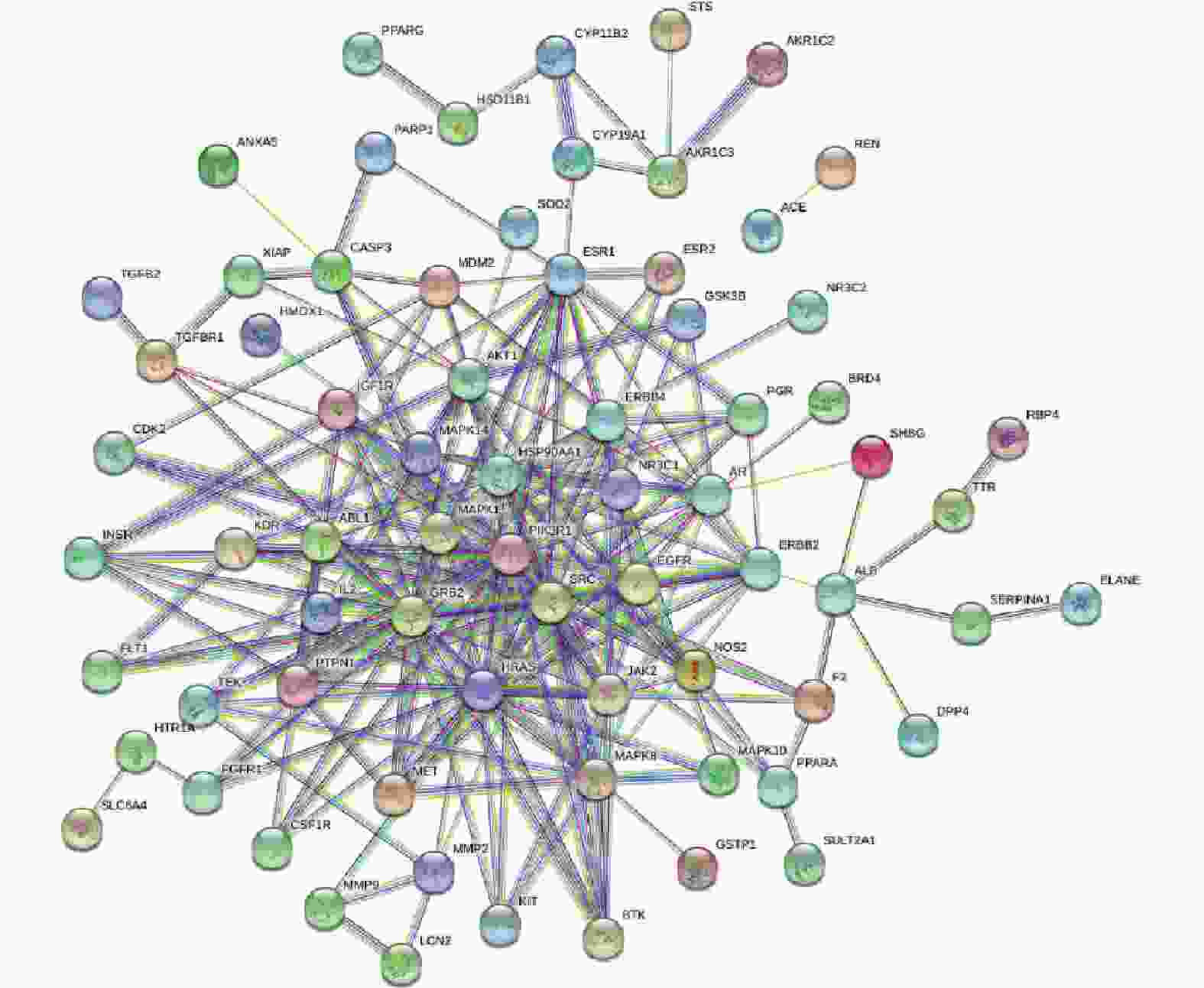
将交集靶点导入String数据库,绘制PPI网络关系图,如图2所示,其中包括网络节点90个,边235条。将共同靶点导入Cytoscape 软件进行核心靶蛋白筛选。如图3所示。综合节点度值及本研究相关度排名靠前的分别为AKT1、ESR1、MMP9、PPARγ、MMP2(见表1)。
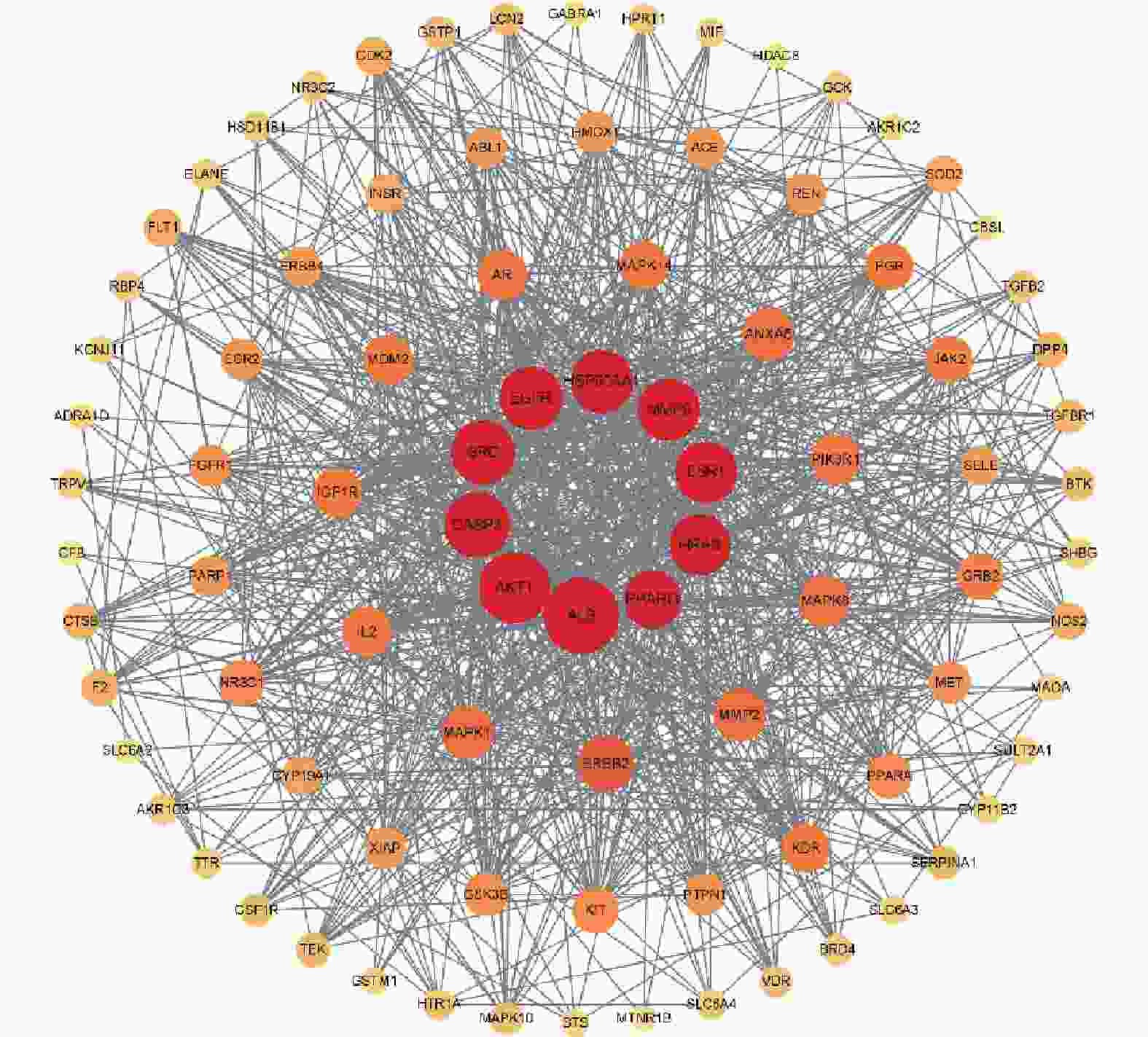
基因名称 节点度值 排名 ALB 66 1 AKT1 60 2 CASP3 53 3 SRC 51 4 EGFR 50 5 HSP90AA1 49 6 MMP9 48 7 ESR1 48 8 HRAS 47 9 PPARγ 43 10 ERBB2 41 11 MMP2 37 12 -
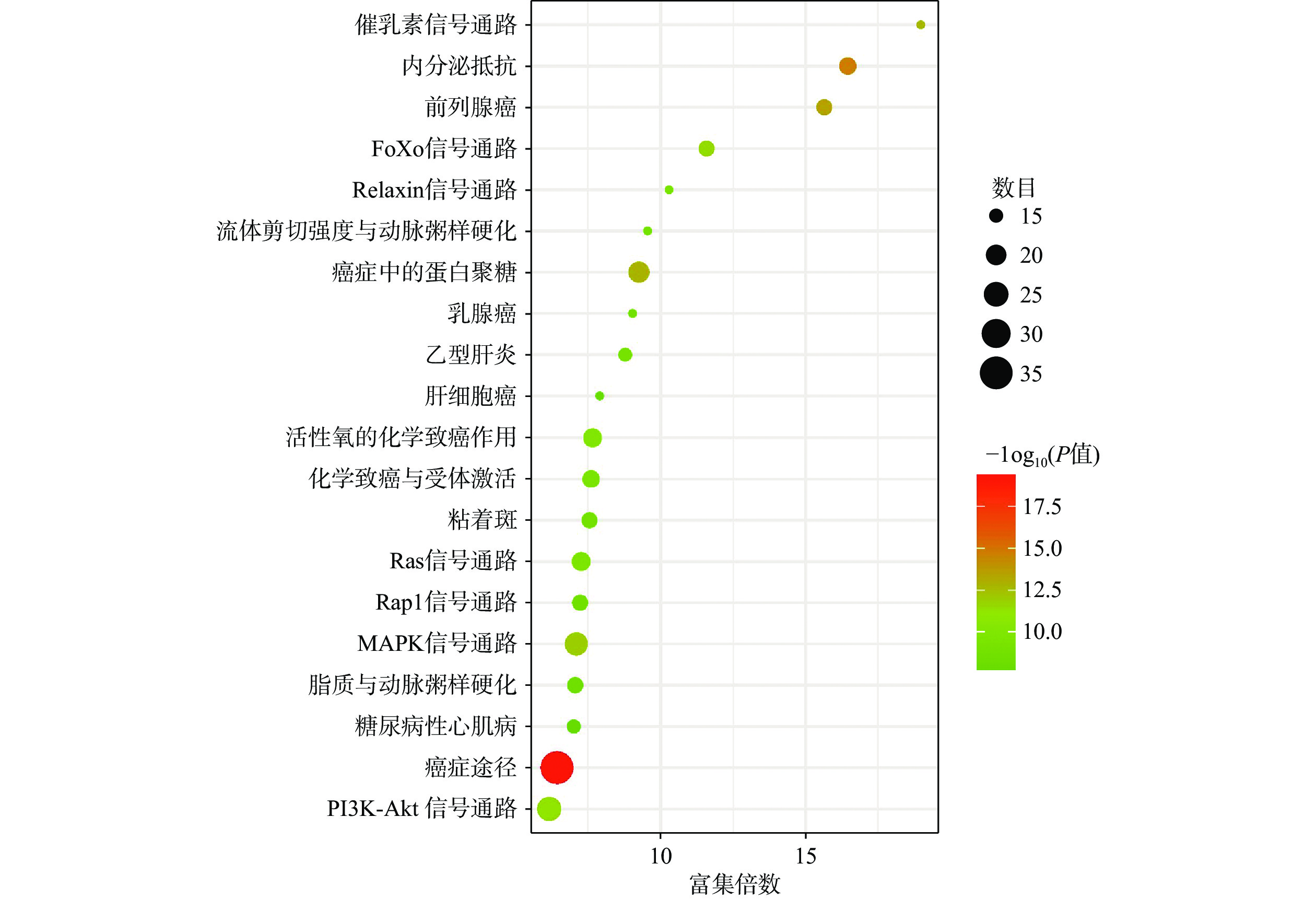
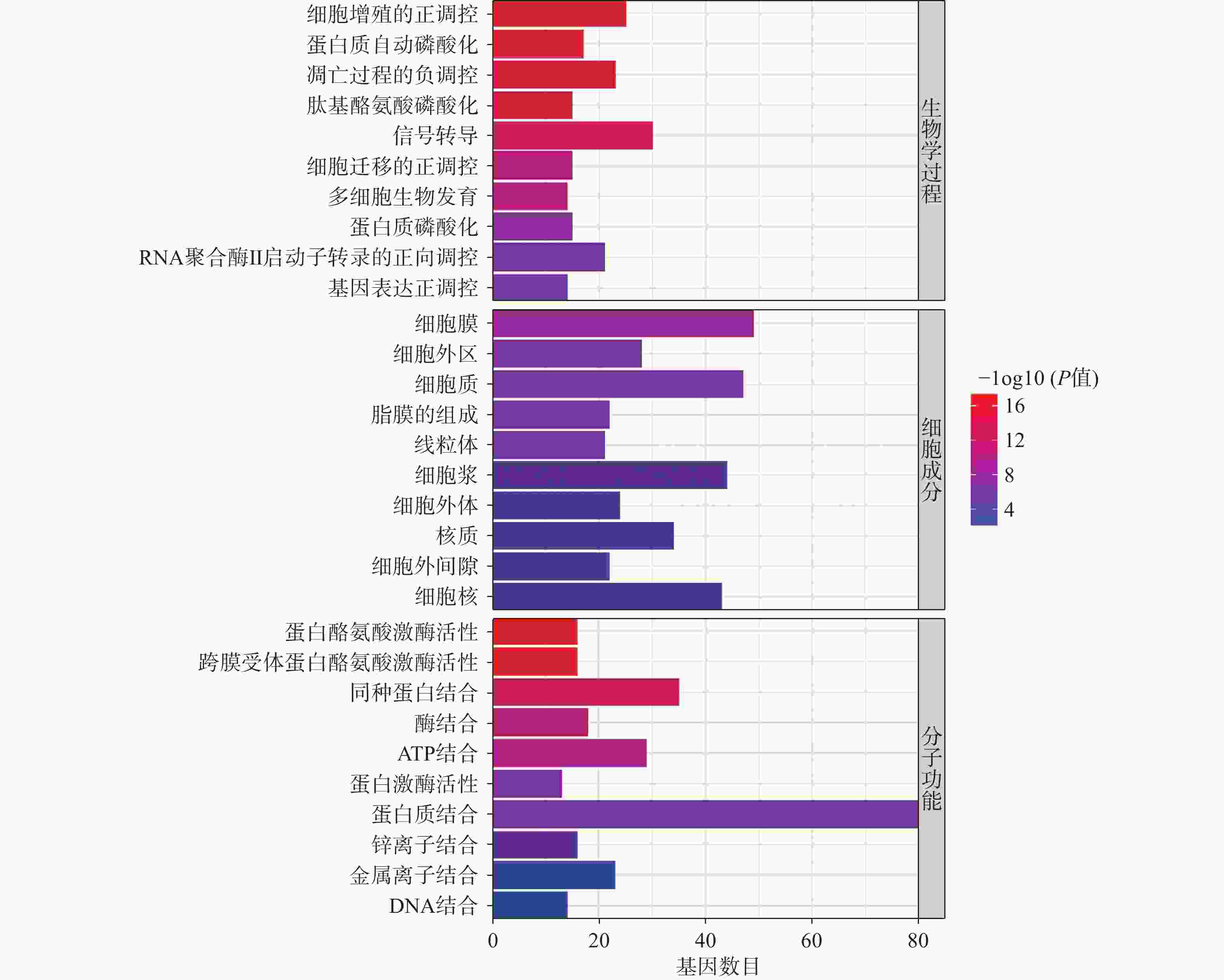
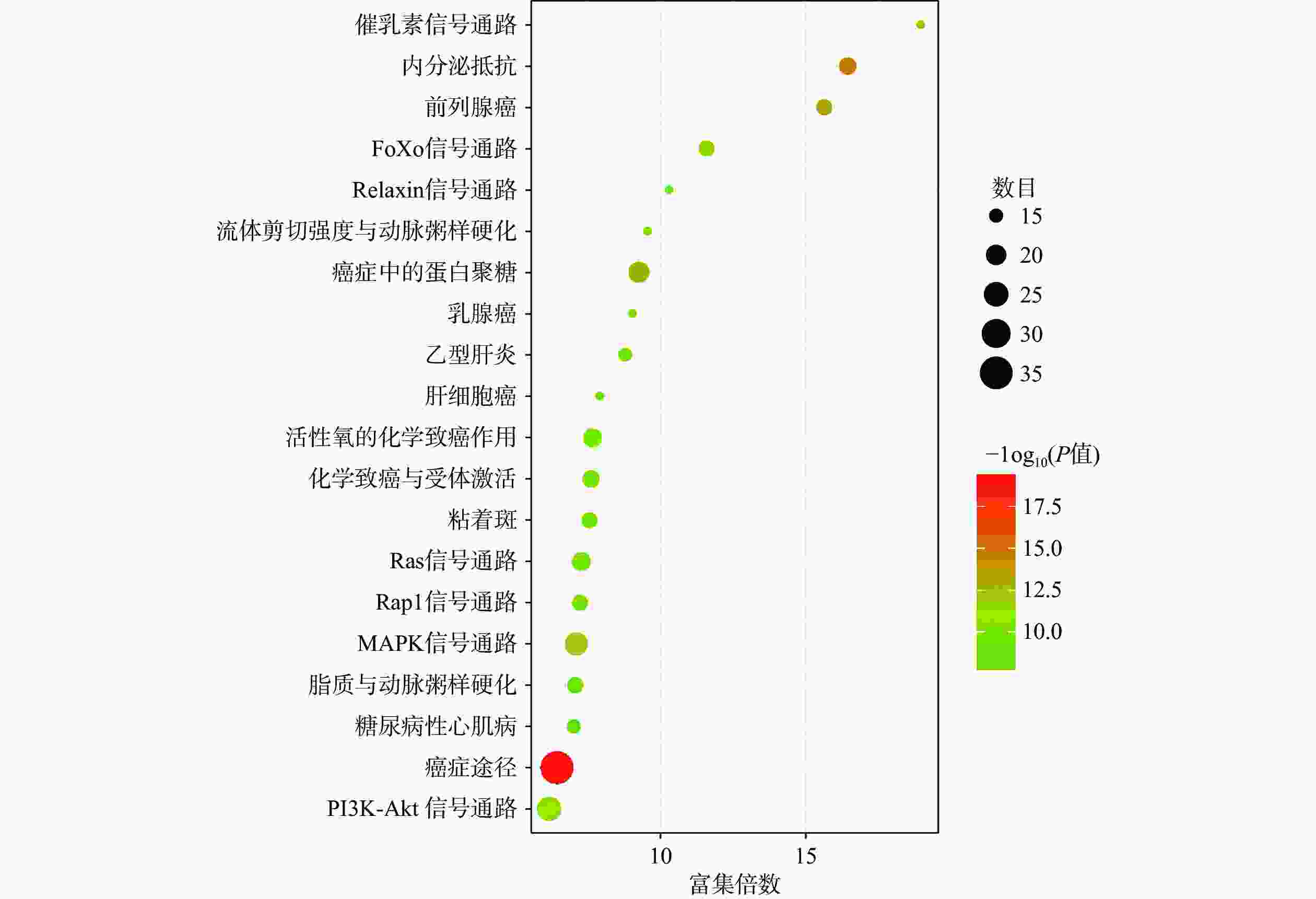
将得到的90个交集靶点导入DAVID数据库进行GO富集分析,富集结果分别根据基因富集程度进行排序,其中BP前10个条目主要涉及细胞增殖调控、蛋白质磷酸化和RNA聚合酶Ⅱ启动子转录的正调控等生物学过程,MF前10个条目主要与蛋白酪氨酸激酶活性、蛋白激酶活性、蛋白结合和酶结合等分子功能有关,CC前10个条目主要在细胞膜、胞质和胞核等部位富集,如图4所示。KEGG富集分析共筛选到162条信号通路,根据基因富集程度排序,前20个条目主要涉及PI3K/Akt、MAPK、Ras、内分泌抵抗等信号通路,如图5所示。
-
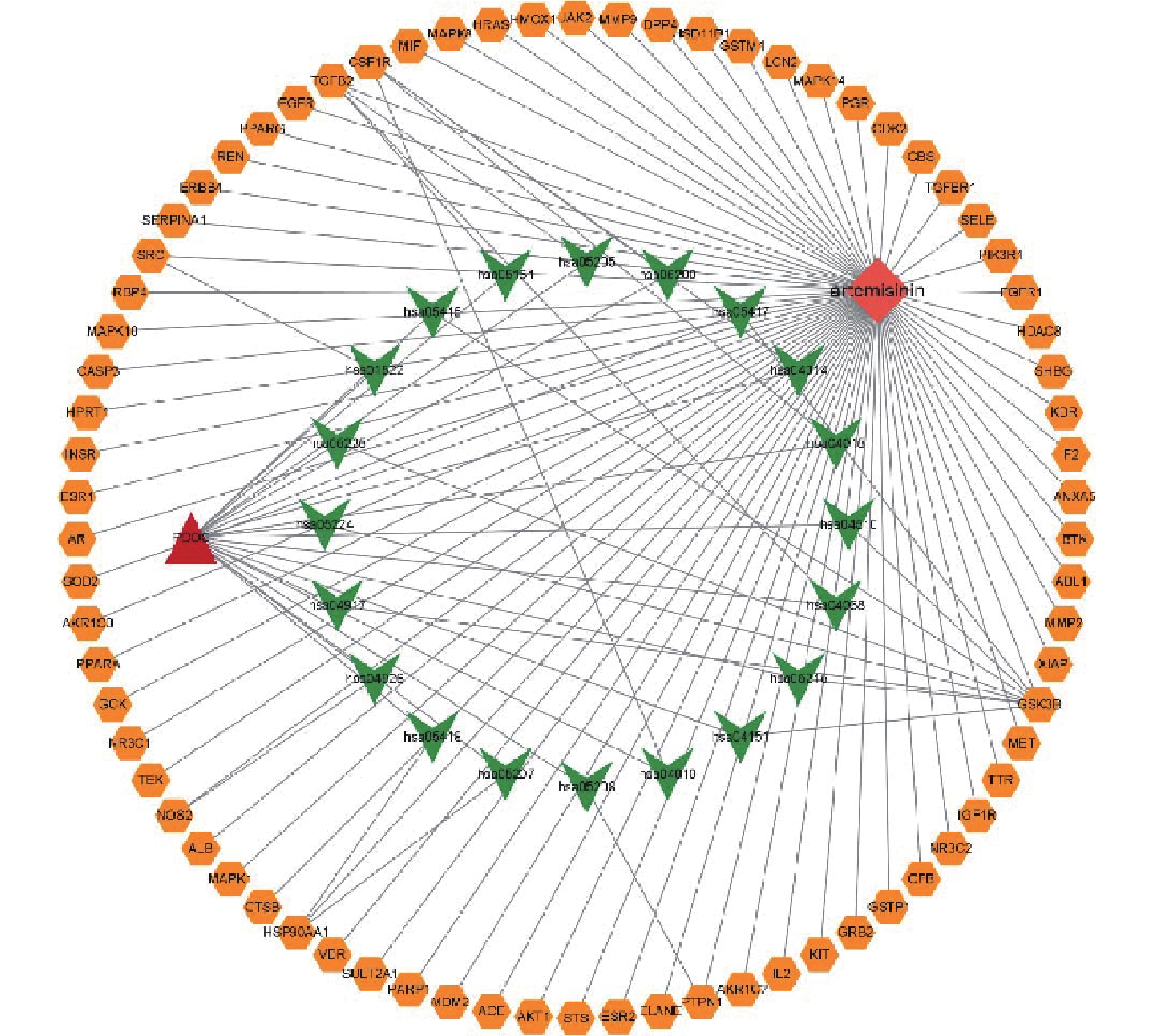
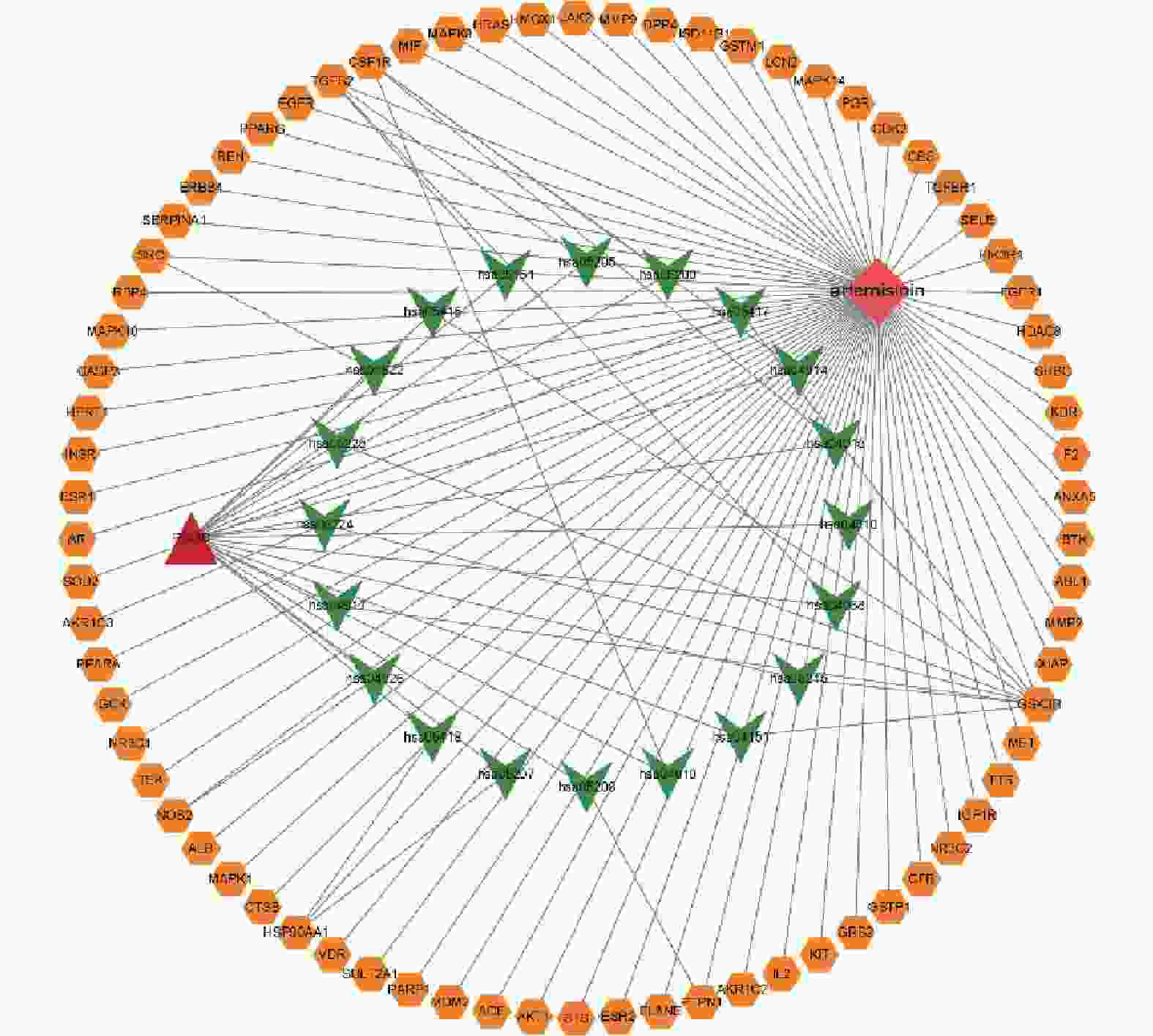
将相关靶点及通路文件导入Cytoscape 3.9.1软件,得到药物、疾病、靶点和通路之间的关系图(见图6)。
-
分子对接结果显示,青蒿素与核心靶蛋白AKT1、MMP9、ESR1、PPARγ、MMP2之间均存在结合位点。青蒿素与核心靶蛋白的最低结合能分数见表2,结合能越低表示结合活性越高,化合物越容易与该靶点结合。其中青蒿素与核心靶蛋白之间的氢键连接可视化情况如图7所示。
化合物 核心靶点 最低结合能(kJ/mol) 结合位点 青蒿素 MMP9 −8.2 ARG-143 AKT1 −7.9 HIS-152 ESR1 −7.9 THR-460 PPARγ −7.7 PRO-426、GLN-430、
LEU-431、PHE-432MMP2 −6.4 HIS-190 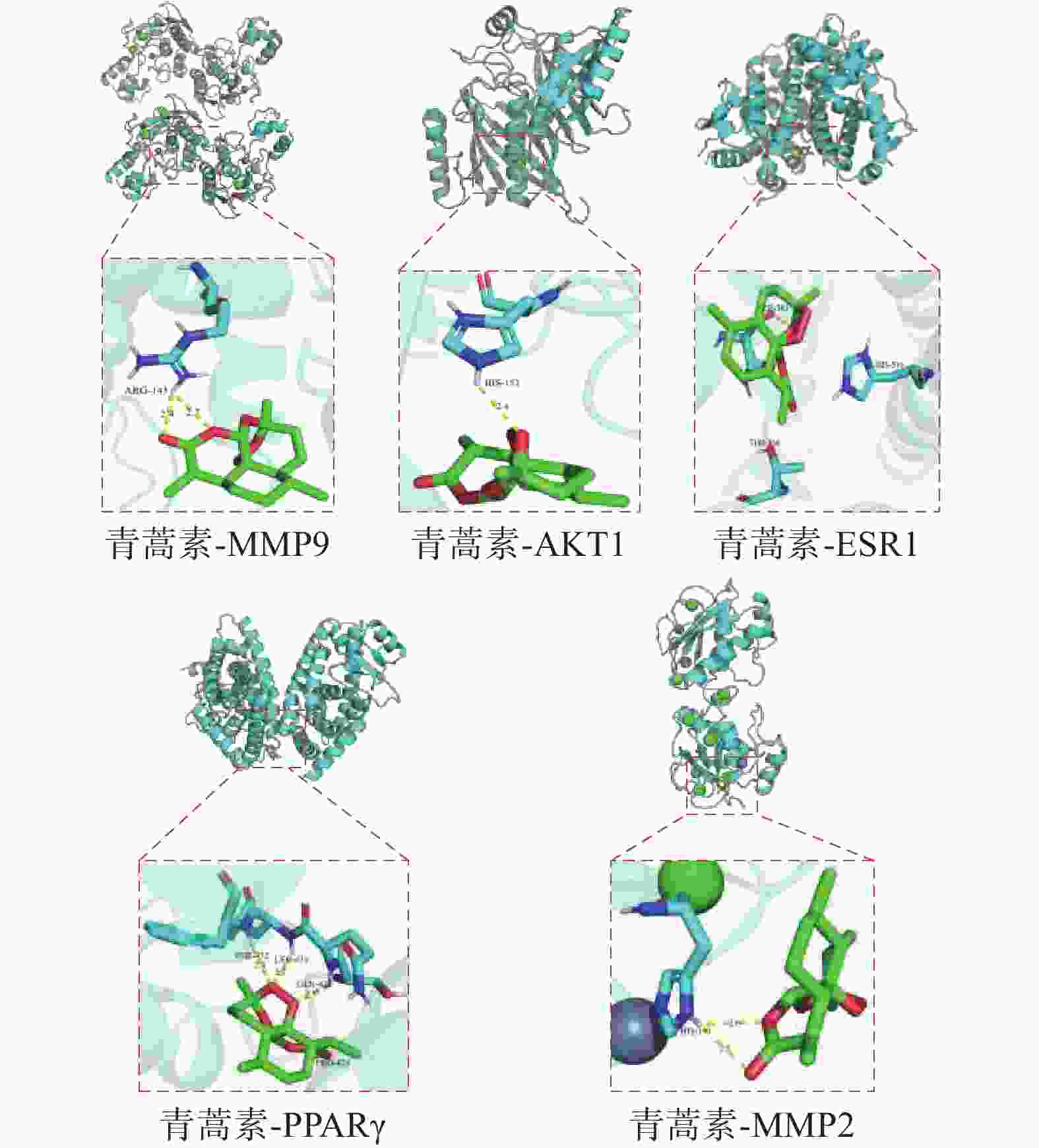
-
PCOS主要通过卵巢病变、机体内分泌紊乱等方式影响女性的生育能力[28]。目前关于PCOS的发病机制和病因尚未具体阐明,多认为是遗传和环境因素相互作用的结果;由于病因机制不明,临床治疗尚无统一方案,多采用对症治疗,如基础生活方式调整,通过控制饮食、增加体育运动以降低体质量和缩小腰围,增加机体胰岛素敏感性,降低胰岛素及雄激素水平,同时辅以相应的药物治疗,以减轻症状。
本研究基于临床发现,PCOS患者多伴有肥胖表现,遂以肥胖与PCOS之间的潜在联系为出发点,同时基于课题组现有天然活性物质进行药物筛选,经文献调研发现青蒿素有抗肥胖效果,继而通过网络药理学方法分析预测青蒿素可能用于治疗PCOS的潜在作用靶点,并探讨其可能用于临床治疗PCOS的可行性。
本研究根据PPI网络拓扑属性分析筛选出核心靶点AKT1、MMP9、ESR1、PPARγ、MMP2等,推测这些可能是青蒿素用于治疗PCOS的潜在作用靶点。有研究表明[29],AKT1在颗粒细胞增殖中起关键作用,而其表达量的高低主要与机体雄激素水平异常有关,这会导致PCOS患者卵巢颗粒细胞正常功能受损。此外,AKT1还具有组织特异性,Song等[30]通过小鼠实验发现脂肪组织中AKT1的选择性抑制可以刺激白色脂肪组织发生褐变,从而可增加机体能量消耗发挥抗肥胖的效果。基质金属蛋白酶(matrix metalloproteinase,MMPs)是一种锌依赖性酶,可由卵巢产生,在卵泡发育和PCOS的发病机制中起重要作用[31];研究发现PCOS女性患者MMPs活性增加,其血液、卵泡液和颗粒细胞中MMP9、MMP2水平升高,高水平的MMPs会通过改变细胞外基质重塑,引起异常卵泡闭锁和卵巢基质组织增加,从而对患者的排卵和生育能力产生不良影响[32]。Barbara等[33]在对正常女性和肥胖女性血清样本中的MMP浓度对比发现,体质量增加可影响女性血清中的MMP浓度。ESR是维持卵巢颗粒细胞分化、卵泡和卵母细胞生长发育以及排卵功能的关键受体[34];ESR1是一种核激素受体,作为转录因子的激活剂发挥作用[35];Schomberg等[36]在ESR基因敲除的小鼠模型中发现ESR基因缺失会导致卵泡发育受阻,以致卵泡闭锁及无排卵现象发生。Artimani等[37]在评估PCOS患者颗粒细胞中ESR基因表达时发现,ESR mRNA的表达显著低于排卵功能正常女性,认为ESR基因的显著减少可作为颗粒细胞成熟缺陷或卵泡发育停滞的指标。ESR1也是一种与线粒体功能相关的基因,研究发现其在肥胖女性脂肪组织中有减少,Zhou等[38]在人类和啮齿动物实验中证实,脂肪组织中ESR1的表达与脂肪量呈负相关,同时ESR-α作用的降低还会损害线粒体功能,促进肥胖增加,破坏机体代谢稳态。PPARγ是一种调节脂肪细胞发育和葡萄糖稳态的核受体,主要在脂肪组织中表达[39];此外在发育阶段的卵巢颗粒细胞中表达,并可受黄体生成素(luteinizing hormone,LH)水平的影响来调节机体雌激素分泌和卵巢功能[40]。此外,Lee等[41]在PCOS患者颗粒细胞中发现PPARγ mRNA表达水平下调。胡卫红等[42]研究发现PPARγ 在PCOS患者的卵巢颗粒细胞的表达异常可能与PCOS的高雄激素血症有关。
GO生物学过程富集分析表明,青蒿素治疗PCOS的生物学功能可能与细胞增殖调控、蛋白质磷酸化和RNA聚合酶Ⅱ启动子转录的正调控等生物过程有关。KEGG通路富集分析表明,青蒿素可能通过作用于PI3K/Akt、MAPK、Ras、癌症等信号通路发挥治疗作用。有研究表明,PI3K/Akt信号通路参与调节细胞增殖分化和迁移,在卵泡发育过程中对卵巢颗粒细胞的生长和凋亡起着关键作用[43],在PCOS患者颗粒细胞中与氧化应激相关的凋亡多伴随PI3K/Akt信号下调[44]。此外,研究表明PI3K/Akt信号通路还可以调节脂肪细胞的脂解与分化,从而参与机体脂质代谢[45]。MAPK信号通路参与调节多种细胞过程,如增殖、分化、转录调控等,且该通路与卵巢颗粒细胞类固醇激素的合成有关[46]。研究发现,在PCOS女性中,异常的MAPK信号传导可导致代谢信号缺陷和卵巢雄激素分泌异常增多[47]。
为进一步探索青蒿素在PCOS治疗中的潜在分子机制,本研究将天然产物青蒿素和5个与PCOS密切相关的核心靶蛋白进行分子对接验证,寻找二者之间存在的最佳结合位点以及评估它们之间的结合能力。验证结果显示,青蒿素与核心靶蛋白之间能够较好结合。
综上所述,本研究采用网络药理学方法分析天然物青蒿素用于治疗PCOS的潜在作用靶点,其机制可能主要涉及PI3K/Akt、MAPK、内分泌抵抗等信号通路。这些信息为后续青蒿素用于治疗PCOS的实验验证提供了重要理论依据。
Exploration on the potential therapeutic mechanism of artemisinin in polycystic ovary syndrome based on network pharmacology and molecular docking technology
doi: 10.12206/j.issn.2097-2024.202209051
- Received Date: 2022-09-20
- Rev Recd Date: 2023-05-20
- Available Online: 2023-12-22
- Publish Date: 2023-12-25
-
Key words:
- artemisinin /
- polycystic ovary syndrome /
- network pharmacology /
- molecular docking
Abstract:
| Citation: | YU Weili, WEI Yifang, YE Zishao, LIU Aifen, WANG Chengniu, ZHANG Lei. Exploration on the potential therapeutic mechanism of artemisinin in polycystic ovary syndrome based on network pharmacology and molecular docking technology[J]. Journal of Pharmaceutical Practice and Service, 2023, 41(12): 714-721. doi: 10.12206/j.issn.2097-2024.202209051

|
 DownLoad:
DownLoad: